
Ethology and animal behaviour are two terms that are now considered synonymous. However, for a long time, they were not, leading to significant disputes in the scientific community and beyond. Let us explore the reasons why.
Since the beginning of the last century, the study of animal and human behaviour using scientific methods has led to the emergence of two contrasting approaches: European ethologists and the so-called behaviourists, primarily American, based on fundamentally different schools of thought and principles. Behaviourists, trained in comparative psychology, firmly believed that humans and animals learn all their behaviours during juvenile development through imitation and learning from conspecifics, parents, and individual experiences. They therefore focused on the environment in which an individual lives as the primary determinant of behaviour.
For ethologists, who come from zoological backgrounds, all this had limited validity, as animals develop a vast repertoire of innate behavioural patterns (instincts) that evolve according to specific developmental programs.
From a philosophical and sociological perspective, the two lines of thought lead to a markedly different vision of the mechanisms of behavioural development, with fundamental implications for humans as well. Are we all born equal, and does the environment shape us differently, or do we carry with us a conditioning experience? In the biological field, it was the “heredity-environment” (nature-nurture) conflict that inflamed tempers, particularly the meaning given to the words “innate” and “instinct,” so dear to ethologists but which behaviourists refused to even acknowledge as drivers and regulators of behaviour.
Renowned physiologists such as Pavlov and Bekhterev, active in the first half of the 20th century, considered behaviour to be generated by simple reactions to external stimuli (reflexes). They argued that complex behavioural sequences were merely chains of reflex responses. They rejected the existence of innate elements in behaviour and dismissed subjective concepts such as attention, will, and individual motivation, asserting that only stimuli and subsequent reflex reactions were objective and demonstrable.
Watson, the founder of behaviourism during the same period, was a strong advocate for the influence of the environment on behaviour, denying any hereditary or genetically transmitted basis (see Note 11).
However, he fully embraced the reflexological views prevalent at the time. From an experimental and methodological perspective, reflexologists and behaviourists can be criticised for their obsessive focus solely on learning processes, even when working with animal models, neglecting phenomena such as appetentia (appetite) and behavioural spontaneity. These aspects are difficult to observe under the experimental conditions they favoured, such as mazes and learning cages, also known as Skinner boxes, named after the psychologist who introduced them into experimental practice. (Image1).

A typical Skinner box. The mouse randomly presses one of the two levers, receiving food from one but nothing from the other. In this way, it learns an association between its behaviour (pressing the correct lever) and a positive consequence (food). After a few uncertainties, it will never make a mistake again.
Under these conditions, all behaviours displayed by the experimental subjects (almost always mice or pigeons) were consequences of the stimuli presented. However, while a hungry pigeon could learn to insert a coin into a vending machine to obtain food, this was not possible if it had two coins at its disposal: in that case, the pigeon’s more specific impulse to rub them together prevailed.
Behaviourists and reflexologists, in their attempt to discover the laws governing behaviour, used a physical-mechanistic approach, overlooking that behaviour operates within a biological-vitalist context, with organisms shaped by the environmental conditions experienced during their natural history. In biology, it is essential to address the purpose of an organ as well as behaviour as part of the phenotype—that is, the adaptive process that generated it and its function—within a distinctly evolutionary framework.
This is precisely the task of ethology, as a comparative study of animal behaviour: examining its phylogenetic dimension. A science with a clear zoological-Darwinian origin, enriched by the ideas of Konrad Lorenz, Niko Tinbergen, and Karl von Frisch, its pioneers in the period before and after the Second World War.
With these masters, the scientific study of behaviour left behind the gloomy Pavlovian laboratories and Skinner’s claustrophobic cages for natural environments, observing behaviour where it has been shaped by external selective pressures, revealing its full range. For them, ethology is the study of instincts, and it is no coincidence that Tinbergen, in 1951, titled his seminal text The Study of Instinct, dedicated precisely to the instinctive, hereditary, and species-specific components of behaviour. This highlights the key difference emphasised between ethology, as the study of the innate components of behaviour, and the behaviourist view, which focused on the learned components as solely responsible for behavioural development in individuals.
But what is behaviour and what are instincts?
The natural behaviour of any organism includes actions such as muscular contractions or their absence, the emission of sounds, light, heat, electricity, the release of secretions and excretions, and colour changes. Many behavioural expressions result from various forms of learning, but just as many—often exclusively—are not learned; instead, they arise from innate, species-specific mechanisms that are often highly resistant or unaffected by previous experiences. For the ethologist, behaviour is part of the phenotype of a given species, representing an integral aspect of its natural history. Therefore, behavioural expressions have predictive value in describing and determining phylogenetic similarities or differences, much like anatomical traits. This perspective is difficult for a behaviourist to accept, as they believe behaviour is shaped by an individual’s personal history rather than by its phyletic and genetic evolution.
But what are instincts according to the ethologist?
Lorenz proposed three distinct concepts for instincts: fixed action patterns (FAP), innate releasing mechanisms (IRM), and imprinting, considering them the observable manifestations of instincts. For Lorenz, FAPs were “the core of instinct“, activated as actions controlled by the central nervous system upon the appearance of environmental stimuli (IRM) that triggered them in a specific and stereotyped manner. For example, at the sight of a sexually mature female, all males of that species display their courtship with identical and specific actions or postures (FAPs), guided by equally specific nerve centers. One of the most iconic FAPs in ethology is the threat display of the male stickleback (Gasterosteus aculeatus) described by Tinbergen: in spring, this small fish stakes out its reproductive territory, and its belly turns bright red. If a conspecific with the same red belly approaches its territory, it immediately approaches and places itself upside down in front of it (the FAP), inducing it to desist and flee (Image 2).

A red-bellied stickleback (Gasterosteus aculeatus) symbolizing sexual maturity and territoriality, and its head-down threat posture.
The intruder’s red belly is the innate trigger stimulus (IRM) that activates the specific motor nerve center, causing it to lie down. If experimentally shown a wax decoy with a red lower half, which does not at all resemble a small fish, it still assumes an upright position (Images 3a and 3b).


Images 3a-3b: Stickleback decoys with red underparts triggering immediate aggression in a territorial stickleback, which instead ignores a very similar one without the spot.
Clear evidence that it is the red spot that triggers the deterrent action. Another similar example of IRM is the red spot on the chest of robins. (Image 4).

Even in the robin, a red spot on the chest of a conspecific stimulates aggressive action in a territorial specimen.
FAPs can combine with other innate behavioural elements to give rise to complex, higher-order actions. This was a fundamental discovery made by Lorenz and Tinbergen while studying egg-retrieval behaviour in the greylag goose (Image 5).

As all ground-nesting birds know, if an egg accidentally rolls out of the nest, they extend their beaks beyond the egg and roll it backward, pushing it with the underside of their beaks, carefully balancing it with lateral movements of their necks to prevent it from slipping. This behaviour consists of two components: if we take the egg away from the goose after it has already started rolling it backward, the neck movement continues in vain, as if it were rolling the egg back toward the nest, due to the activation of the respective nerve center that completes its motor command. However, the lateral balancing movements are suppressed: the neck moves in a straight line toward the nest. This movement, once set in motion, continues even without the need for external stimuli (the egg) and is the FAP. Lateral corrective movements are orientation actions or taxis, also innate but which stop when the triggering stimuli cease (the egg slipping away). Lorenz called FAPs “innate” to emphasize that the ability of a given species to produce them in a stereotyped way did not depend on learning but was performed perfectly without any previous experience.
However, instinct is much more than a mere collection of movements or postures (FAPs) and specific reactive sensitivities (IRMs). It includes the central control structures that allow a species to behave in the manner best suited to its environment. All of this is a response to evolutionary processes that have occurred within the natural history of a species, and therefore within its phylogeny.
Imprinting is still an innate behaviour, the characteristics of which are the fruit of the genius of Lorenz, who discovered them while studying the behaviour of birds, especially ducks. Their young are induced to consider the first being they encounter upon hatching as their “mother,” establishing a specific and lasting relationship with it.
The term “ethology” is linked to the study of instinctual and unlearned behaviours, considering them the primary and defining elements of behaviour. These phenomena, called Fixed Action Patterns (FAPs), undergo adaptive changes that can sometimes be observed in species related through evolutionary kinship. A well-known example of FAPs is found in a family of insects (Family Empididae, Hymenoptera) whose males present females with ritual gifts before mating. In two of these species, males capture an insect and offer it to the female, calming her aggression. In two others, males wrap their prey in a balloon-shaped cocoon, spinning it with their tarsal glands, which keeps the female occupied for longer. Another species presents a balloon-shaped cocoon containing only a petal or a small leaf, making it a purely ritual gift. Finally, males of Hilaria sartor spin the balloon and use it to present themselves to the female as a sexual signal (Image 6).
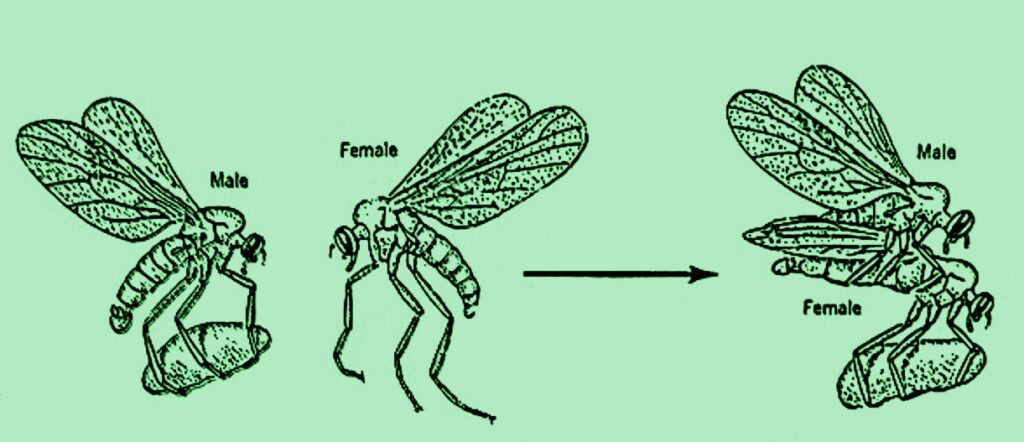
This last behaviour can only be understood in light of what other closely related species do.
Towards the encounter between two different schools
However, the very definition of behaviour we gave above cannot ignore the elements learned throughout an individual’s life. Lorenz himself knew this well when he recounted how, on the path leading to the goose pond, a tree branch had fallen. A small family of shrews walking along the path jumped over it, and so did the young ones following in single file. Once the branch was removed after a few days, the shrews that reached that point still jumped as if it were still there, driven by a stereotyped reaction they had quickly learned. It therefore seemed both sterile and unproductive to continue pitting innate versus learned, nature versus nurture, heredity versus environment, as the foundation of behaviour—a view ethologists and behaviourists had championed for years. It had been a long cultural and academic battle between the opposing shores of the Atlantic, which seemed to end when, in 1973, Lorenz, Tinbergen, and von Frisch were awarded the Nobel Prize for Medicine and Physiology for their “ethological” studies.
Vissi in “prima fila” quel momento, ero infatti in Olanda, patria di Tinbergen, a studiare Etologia e con un po’ di fortuna, le mie ricerche sui cambiamenti di colorazione dei pesci Ciclidi furono menzionate nel libro che gli zoologi olandesi dedicarono a Tinbergen in quella occasione. I cambiamenti di colorazione dei Ciclidi sono elementi istintuali del comportamento, in rapporto a situazioni sociali legate all’aggressività territoriale e riproduttiva, del tutto innati. Erano innate anche le direzioni di migrazione dei giovani pettirossi che studiavo a Seewiesen in Baviera nell’istituto Max Planck diretto da Lorenz, qualche anno dopo. Ma guarda un po’ la bussola stellare con cui orientavano il loro volo notturno se la dovevano apprendere durante le prime settimane di vita… e così i miei piccioni viaggiatori apprendevano la loro mappa di navigazione dagli stimoli ambientali percepiti nel luogo in cui erano allevati.
I was in the front row at that time, having been in Holland, Tinbergen’s homeland, studying ethology. With a bit of luck, my research on the color changes of Cichlid fish was mentioned in the book that Dutch zoologists dedicated to Tinbergen on that occasion. Cichlid color changes are instinctive behaviours related to social situations involving territorial and reproductive aggression, and are entirely innate. The migration directions of the young robins I studied at Seewiesen in Bavaria at the Max Planck Institute, directed by Lorenz a few years later, were also innate. And just imagine, the star compass with which they oriented their nocturnal flight had to be learned during the first weeks of life… so my carrier pigeons learned their navigation map from the environmental stimuli perceived in the place where they were raised.
Instincts alone did not explain the multifaceted complexity of animal and human behaviour. After 1973, scientific circles gradually embraced a new synthesis in which behaviour was recognized as the interaction between heredity and elements learned by the individual. Lorenz himself spoke of “closed” or “open” behavioural programs: in the former, the innate element emerges predominantly or exclusively; in the latter, the programs require the acquisition of learned elements to reach maturity. Perhaps more compelling is the view of behaviour as a continuum, with innate behavioural modules resistant to external conditioning at one extreme, and those dependent on learning at the other. For a given species, the evolutionary equilibrium point depends on the conditions most favorable to its survival and reproductive success. Natural selection thus favors the optimal combination of instinctive and learned elements in behaviour.
Culturally, it was challenging to introduce such a vision of behaviour, but a unity of goals and methods was achieved for the current model of study and a better understanding. New disciplines emerged from this synthesis, such as neuroethology, sociobiology, and behavioural ecology. The divisive perspective between ethologists and behaviourists soon dissolved, losing its significance, making ethology and animal behaviour synonymous terms.
_____________________
“Give me a dozen healthy infants, well formed, and my own specified world to bring them up in and I’ll guarantee to take any one at random and train him to become any type of specialist I might select – doctor, lawyer, merchant, chief and yes, ever beggar man and thief, regardless of his talents, peculiarities, tendencies, abilities, vocations, and race of his ancestors.”
J. B. Watson 1930. Behaviorism. W.W. Norton & Company, Nev York. Pag. 104.
Credits
Author: N. Emilio Baldaccini Former Professor of Ethology and Conservation of Zoocenotic Resources at the University of Pisa. He has published over 300 scientific papers in national and international journals. Actively engaged in scientific education, he is also a co-author of academic textbooks on Ethology, General and Systematic Zoology, and Comparative Anatomy.
Translated by Maria Antonietta Sessa


{kind=link}
{kind=link}
{kind=link}
{kind=link}