

Il campo geomagnetico come fonte di informazioni direzionali
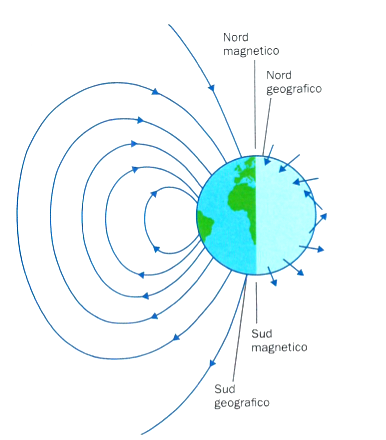
La Terra, per le sue caratteristiche costitutive, è assimilabile ad una enorme calamita. Viviamo così, volenti o nolenti, immersi nelle linee di forza del suo campo magnetico. Queste avvolgono il nostro pianeta partendo da un punto prossimo al polo sud geografico, per rientrare in vicinanza di quello nord, andando a costituire i poli terrestri magnetici. Nel loro percorso, le linee di campo assumono angoli di inclinazione rispetto all’orizzontale continuamente variabili, scorrendo parallele alla superficie terrestre solo nei pressi dell’equatore magnetico (Fig. 1).
L’intensità del campo, inoltre, varia in modo significativo: decresce da 60.000 a 30.000 nT (Tesla è l’unità di misura magnetica) andando dai poli all’equatore magnetici. Inoltre l’andamento delle linee di forza è complicato da deviazioni locali dipendenti dalla composizione dei materiali della crosta terrestre. Rocce ferrose le distorcono infatti notevolmente e, tanto per fare un esempio, chi va per mare sa bene che le nostre bussole non dicono il vero nelle vicinanze dell’isola d’Elba, per i depositi ferrosi che ne costituiscono il substrato.
Le nostre bussole non fanno altro che allineare i lori aghi magnetizzati alle linee del campo, indicando così invariabilmente il nord magnetico. Ogni animale che fosse sensibile al campo geomagnetico sarebbe allora in grado di ricavare da esso informazioni direzionali: avere quindi una bussola magnetica.
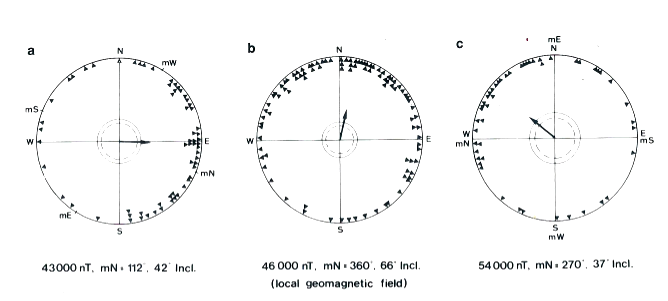
Che gli animali siano sensibili a stimoli orientanti magnetici è ampiamente fuor di dubbio, basta osservare nella figura 2 cosa accade al comportamento di orientamento di uccelli passeriformi, se ruotiamo la posizione del polo nord magnetico da essi percepito: in primavera un pettirosso in migrazione si orienta verso nord, la sua giusta direzione di volo (2b). Se per mezzo di spirali di Helmholtz, poste attorno alla sua gabbia, ruotiamo verso ESE il nord magnetico, anche le sue scelte direzionali deviano in tale direzione (2a). Lo stesso accade se il nord è posto in ovest (2c). Egualmente un colombo viaggiatore può avere difficoltà ad orientarsi verso casa a cielo coperto da luoghi a forte deviazione delle linee di campo, come ho osservato in lanci dall’area vulcanica viterbese, in modo analogo a quanto fatto dai colleghi della Cornell University (NY) da un luogo detto, guarda caso, Iron Mine.
Se la risposta comportamentale degli uccelli non lascia dubbi sulla loro sensibilità magnetica, non mancano tuttavia alcune perplessità in merito, dovute soprattutto a due motivi:
– nessuno è riuscito a condizionare gli uccelli a stimoli magnetici;
– non sappiamo quale sia il recettore responsabile della sensibilità magnetica, e questo per nessuna specie.
Dagli esperimenti di tipo pavloviano sappiamo che un uccello può essere condizionato da stimoli sonori, visivi o elettrici, a compiere un certo movimento o azione, ma nessuno di essi ha mai risposto a stimolazioni magnetiche.
Similmente, la ricerca di un “recettore magnetico” ha a lungo impegnato fisiologi ed anatomici, finora senza successo apprezzabile. Le teorie sono molte ma i risultati sperimentali appena incoraggianti e nessuno certo. Oggi l’ipotesi più promettente si basa su di un meccanismo mediato da proteine (criptocromi) presenti nella retina di uccelli e mammiferi. Tuttavia tra i vari subtipi di criptocromi presenti in parti differenti della retina, un sicuro candidato come proteina primaria per la magneto recezione non è ancora stata individuata. Ancora più incerta la via nervosa che dovrebbe portare l’informazione retinica ai centri decisionali dell’encefalo.
Ma allora esiste o no una bussola magnetica? E quali sono le sue caratteristiche funzionali? Anche se sole, luna e stelle sono le mie preferite fonti di informazione direzionale, non posso certo negare il ruolo del geomagnetismo come elemento bussolare primario ed innato negli animali, non necessitando di essere appreso e funzionando addirittura come sistema di calibrazione delle altre bussole appena citate.
Il problema di come gli stimoli magnetici possano essere letti e usati a fini bussolari è stato risolto da Wolfang e Rosita Wiltschko dell’Università di Francoforte nel 1972 (The magnetic compass of European robin. Science 172: 62-64). I loro esperimenti sul pettirosso hanno evidenziato due fondamentali differenze rispetto alle bussole che noi usiamo. Primo fatto è che il range di funzionamento della bussola è sintonizzato sulla intensità del campo in quel punto. Un cambiamento repentino di intensità blocca la lettura della bussola. Un migratore deve quindi adattarsi alle intensità di campo man mano incontrate lungo il viaggio, un fatto che non è immediato ma richiede tempo (2-3 giorni). Secondo, se le nostre bussole sono “a polarità” leggendo la direzione del campo che è sempre verso nord, quella degli uccelli è “ad inclinazione” essendo attenta all’angolo che le linee di campo fanno con la superficie della terra in quel punto.
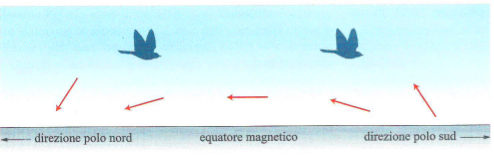
Il parametro inclinazione varia angolarmente tra 90° all’equatore magnetico e 0° ai poli, in quanto le linee di campo sono orizzontali e rispettivamente verticali nei due punti. Inoltre il vettore che lo rappresenta è diretto verso l’alto nell’emisfero australe e verso il basso in quello boreale (confronta con le figure 1 e 3). In accordo a tale parametro gli uccelli non distinguono tra nord e sud, ma piuttosto tra “verso un polo” o “verso l’equatore” magnetici.
Il parametro vettoriale inclinazione si presenta dunque variamente orientato rispetto alla superficie terrestre, ossia forma un certo angolo con il vettore gravità. Vediamo allora nella figura 4 come i Wiltschko hanno ipotizzato il funzionamento della bussola magnetica nel pettirosso: nell’emisfero boreale esso riconosce come “verso nord” la direzione in cui i vettori inclinazione e gravità formano l’angolo α più acuto (4a). Riconosce così la rotta da tenere in primavera per riconquistare le aree di nidificazione europee. Invertendo sperimentalmente le componenti orizzontale e verticale del vettore inclinazione, non si produce effetto sull’orientamento del pettirosso essendo il minor angolo nella solita direzione (4b). Se si inverte la sola componente verticale (4c), o quella orizzontale (4d), si riproduce la situazione dell’emisfero australe, e le preferenze direzionali del pettirosso si invertono, indicando la direzione “verso l’equatore” che il pettirosso percorre in autunno.
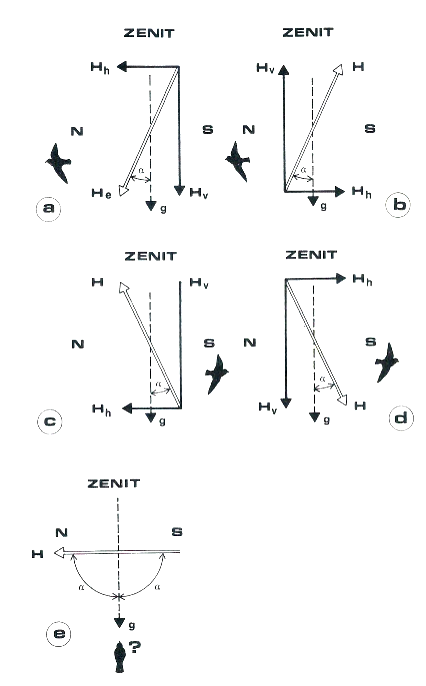
Nelle condizioni equatoriali in cui il vettore inclinazione è parallelo alla superficie e quindi è perpendicolare alla gravità, si formano due angoli di 90°, in tal caso il pettirosso non sa prendere alcuna direzione (4e). Il pettirosso non migra oltre l’equatore, rimanendo a svernare nell’area mediterranea, ma per i migratori transequatoriali che dovrebbero usare lo stesso tipo di bussola, come possono gestire il passaggio nelle aree periequatoriali? Gli stessi Wiltschko ammettono che dal punto di vista bussolare questo “is still an open question”. Per beccafichi, luì e balie nere il passaggio ad una bussola stellare (in quanto migratori notturni) è forse la più facile delle ipotesi, anche se non tale in condizioni di cielo coperto…
La mancanza di modelli alternativi di funzionamento di una bussola magnetica ha fatto si che quello proposto dai Wiltschko sia stato generalmente accettato per gli uccelli, ma come forse traspare da queste righe molti sono ancora gli interrogativi a cui rispondere.
Crediti
Autore: N. Emilio Baldaccini. Già Professore Ordinario di Etologia e di Conservazione delle risorse Zoocenotiche dell’Università di Pisa. Autore di oltre 300 memorie scientifiche su riviste internazionali e nazionali. Svolge attività di divulgazione scientifica. E’ coautore di testi universitari di Etologia, Zoologia Generale e Sistematica, Anatomia Comparata.



{kind=link}
{kind=link}
{kind=link}
{kind=link}